News & Articles
Skeletal Muscle Function Modeled as a Mechanical Linear Electro-Active Actuator/Motor
From Greg Mack
Let’s take a look at some definitions:
Muscle (n.) – a specialized biological tissue system that contains cells capable of generating force.
Mechanical (adj.) – of or relating to machinery or tools, caused by, resulting from, or relating to a process that involves a purely physical as opposed to a chemical or biological change or process.
Linear (n.) – involving measurement in one dimension only; pertaining to length:
Electro-Active (adj.) – (of living tissue) exhibiting electrical activity or responsive to electrical stimuli
Actuator (n.) – a mechanical device for moving or controlling a mechanism or system. It is operated by a source (or several sources) of energy, usually in the form of an electric current, hydraulic fluid or pneumatic pressure, and converts that energy into some kind of motion.
When in-vivo skeletal muscle is activated electrically by the nervous system it generates an internal force via the contraction of its cells. At the most basic level of analysis a muscle produces a resultant vector that acts along a straight line on the skeletal bones (which will be referred to as levers), by way of the tendons and aponeuroses physically attached to the muscle fibers. The muscles and the levers they act upon relate to each other via the synovial joints as an interdependent, interconnected, and interacting system. The joints possess geometries via its architecture produce axes that joints related levers (skeletal bones). This dynamic capability allows relative motion and position maintenance between the various levers of the body. This is a mechanical process for controlling the skeletal system. This control is constructed by the cooperative effort of the anatomically named muscles.
Given this description muscle is an analogous counterpart as a biological model of a human
engineered “Mechanical Linear Electro-Active Actuator”. Understanding the Actuator analogy has important implications for interpreting observed motion and position control under conditions and reaching conclusions that dysfunctions or inefficiencies are occurring in the human musculoskeletal system.
“A linear actuator is an electric motor that has had its stator and rotor “unrolled” so that instead of producing a torque (rotation) it produces a linear force along its length.” Wikipedia
“These concepts demonstrate the elegant manner in which muscle acts as a biologically regenerating linear motor.” Lieber, R.L, “Skeletal Muscle is a Biological Example of a Linear Electro-Active Actuator”, SPIE’s 6th Annual International Symposium on Smart Structures and Materials, 1-5, March 1999
Defining Skeletal Muscle and Its Properties
Defining the boundary conditions of what is to be called “muscle” clarifies and supports the understanding of the Model Mechanical Linear Electro-Active Actuator/Motor and leveraging the Model Mechanical Non-Linear Spring paradigm presented. Therefore, it is necessary to eliminate some of the components of the actual in-vivo anatomical material infrastructure (i.e. blood vessels, nerve cells, lymphatic tissues, bursae) in the description that do not directly support the development of these conceptual models.
Skeletal muscle is a particularly remarkable tissue. As a unique tissue system, it is set within, and integral to, the body’s diverse material infrastructure. Delineating the materials that constitute whole muscle is important in order to develop meaningful boundary conditions that constrain the study to an understanding of the environment in which the muscle system works as a mechanical apparatus acting under forces within a lever system. Whole skeletal muscle includes its myofibrils and the web-like scaffold of various forms of collagen (forming tendons, aponeuroses, myomesium, etc.).
Therefore, the tissues being discussed regarding muscle are those that form a tendon and its attachment to a bone, through to the tendon’s material transition into the myofibril network, the myofibrils, the connective tissue within and around the myofibrils (perimysium, endomysium, etc.), to the myofibril’s transition to the other side tendon, and that tendon’s attachment to another bone. (See Illustration Below).

As described skeletal muscle tissue is constructed from various types of cellular material. Some of these cells constitute the active force generating elements (the sarcomeres which constitute the myofibrils). Other cells constitute the passive connective tissue (tendons, endomysium, epimysium), which serve as scaffolding and/or containment for the active contractile elements. In partnership the active and passive tissues are observed as whole muscle that is given an anatomical name i.e. biceps brachii or pectoralis major. Hundreds of muscles have been identified and given names, each one possessing a unique architecture and relationship within the skeletal lever system. A Latin or Greek root base appellation is provided typically based on a muscle’s configuration within the system. (shape, attachment arrangement, function, and/or location on the skeleton (i.e. Triceps Medial Head, Anterior Tibialis, Extensor Digitorum).
“Each muscle of the body has a general structure that includes various connective tissue components that give the muscle shape and organization.” Macintosh, Gardiner, McComas “Skeletal Muscle: Form and Function”, 2006, Page 3
The Viscoelastic Property of Skeletal Muscle & Modeling it as a Mechanical Non-Linear Spring
Muscle tissue consists of approximately 75% water by weight. The remainder of the tissue is generally considered an amorphous polymeric composite material continuum. Therefore, whole skeletal muscle when viewed from a material perspective, has been referred to as possessing both viscous and elastic behavioral properties.
“As an engineering material, muscles fall into the category of substances that exhibit time-dependent, non-linear viscoelastic behavior.” Schneck, D.J., Mechanics of Muscle: Second Edition, Pg. 51
“Mechanically, the muscle-tendon unit functions as a composite viscoelastic structure.” Magee, Zachazewski, Quillen – Scientific Foundations and Principles of Practice in Musculoskeletal Rehabilitation 2007, Pg. 102
“… collagen is a viscoelastic material; that is, under rapid loading it behaves elastically, while under gradual loading it is viscous and can deform plastically.” Siff, Mel, C., “Supertraining”, 2003, Pg. 43
Viscoelasticity (n.) – the property of a substance of exhibiting both elastic and viscous behavior, the application of stress causing temporary deformation if the stress is quickly removed but permanent deformation if it is maintained.
As a behavioral property of muscle, it is vital to understand why the neologism “viscoelastic” has been ascribed to muscle action during human movement and position maintenance. This will also have implications for interpreting observations of muscle behavior under a variety of conditions. Viscoelastic stems from two separate words: viscosity and elasticity.
Viscosity (n.) – A quantity expressing the magnitude of such friction, as measured by the force per unit area resisting a flow in which parallel layers, unit distance apart, have unit speed relative to one another. The state of being thick, sticky, and semi-fluid in consistency, due to internal molecular friction. Generally identified with the flow characteristics of a fluid.
“Since Sir Isaac Newton originated the concept of viscosity, the study of liquids with strain rate dependent viscosity is also often called Non-Newtonian fluid mechanics.” Wikipedia
“Viscosity is the principal means by which energy is dissipated in fluid motion, typically as heat.” Knowledgerush.com
Viscous (adj.) – having a thick, sticky consistency between solid and liquid.
Elasticity (n.) – the property of a substance that makes it possible to change its length, volume, or shape in direct response to an external force and to recover its original form upon the removal of that force. Generally associated with changes in the length of a solid or semisolid material.
“In physics and mechanical engineering, the theory of elasticity describes how a solid object moves and deforms in response to external stress.” Knowledgerush.com
Elastic (adj.) – (of an object or material) able to resume its normal shape spontaneously after contraction, dilatation, or distortion.
Considering these definitions, the neologism ‘viscoelastic’ requires additional analysis in order to grasp its historical context as a descriptive term applied to muscle behavior. The physical concepts of viscosity and elasticity are generally applied to, and descriptive of, non-biological materials under forces under varying temporal intervals and standardized conditions. Each possess specific measurement with units. Viscoelasticity attempts to capture both acute and chronic time dependent state changes and reflects a material that is capable of both evanescent and permanent behavior.
“By contrast, elastic and viscous (or intermediate, viscoelastic) behavior is relevant at short times (transient behavior).” Wikipedia
Whether the neologism is combining the adjective forms, the noun forms, or mixing the adjective and noun forms, conceptually it attempts to transfer the physics of viscosity and elasticity, which occur in the intrinsic properties of passive non-biological solids and liquids reacting under forces and fluids subject to flow, to that of empirically observed behaviors of active biological materials, like muscle, which can alter the nature of its material continuum via contraction, under forces.
“Human and animal tissues are generally called elastic if one wishes to emphasize their ability to return to a unique configuration when all the external load is removed.” Fung, Y.C., “A First Course in Continuum Mechanics”, 3rd Edition, 1994, Pg. 193
“The viscoelastic behavior of muscle is more complex because of the contractile components found within muscle; however, several of the non-contractile collagenous components exhibit typical viscoelastic properties. The elastic property implies that deformation of a change in length is directly proportional to the load … Viscous properties are characterized by time and rate change dependency.” Magee, Zachazewski, Quillen – Scientific Foundations and Principles of Practice in Musculoskeletal Rehabilitation 2007, Pg. 102
(The previous two quotes create a possibility that the modern notion that whole active muscle possesses viscoelastic properties, in the most formal sense of flow and length change, can be challenged.)
The change attributes of viscoelasticity for a force applied to a solid or fluid material are described by the concepts of hysteresis, stress-relaxation and creep.
Creep (v.) – to undergo slow plastic deformation.
Hysteresis (n.) – the lag in a variable property of a system with respect to the effect producing it as this effect varies. The phenomenon exhibited by a system, often a ferromagnetic or imperfectly elastic material, in which the reaction of the system to changes is dependent upon its past reactions to change.
Stress-Relaxation (n.) – the time-dependent decrease in stress under sustained strain.
Modeling for establishing the mechanical properties of viscoelasticity are represented by an idealized “hookean” linear spring and a hydraulic cylinder (referred to as a dashpot or damper).
Hookean (adj.) (of a material) For which Hooke’s law is a
useful approximation; linear–elastic.
www.yourdictionary.com
Hookean Behavior is the behavior of a perfectly elastic body.
Hooke’s law – the stress applied to stretch or compress a body is proportional to the strain or change in length thus produced.
“A linear spring is supposed to produce instantaneously a deformation proportional to the load.” Fung, Y.C., “Biomechanics – Mechanical Properties of Living Tissues”, 1993, Pg. 42
“The hookean body. The perfect spring provides a model for elastic behavior.”
Magee, Zachazewski, Quillen – Scientific Foundations and Principles of Practice in Musculoskeletal Rehabilitation 2007, Pg. 102
“Masses and springs are energy storage elements. A mass stores kinetic energy, and a spring stores potential energy. The damper is a dissipative element. It uses energy and converts heat.” Norton, Robert L. “Design of Machinery: An introduction to the synthesis and analysis of mechanisms and machines”, 4th Edition, 2008, Pg. 751.
Linear (n.) – Mathematics – able to be represented by a straight line on a graph; involving or exhibiting directly proportional change in two related quantities.
Spring (n.) – An elastic body or device that deforms when a force acts on it and then recovers its original shape when that force is removed.
Viscosity is visualized by analogy to a hydraulic cylinder and represented symbolically as a dashpot or damper. A dashpot is often used to represent the shock absorbing property of a material in contrast to the representation of flow.
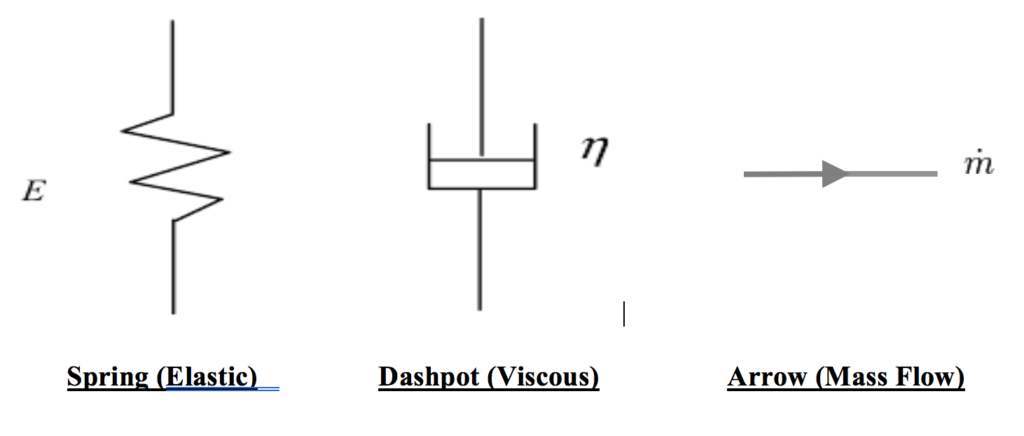
“The newtonian body. A model for viscous behavior is provided by a dashpot or hydraulic cylinder containing a viscous fluid.” Magee, Zachazewski, Quillen – Scientific Foundations and Principles of Practice in Musculoskeletal Rehabilitation 2007, Page 102
“A dashpot is supposed to produce a velocity proportional to the load at any instant.” Fung, Y.C., “Biomechanics – Mechanical Properties of Living Tissues”, 1993, Page 42
Mass Flow (n.) “The mass of a given substance that moves through a given conduit or area per specified unit of time.” www.businessdictionary.com
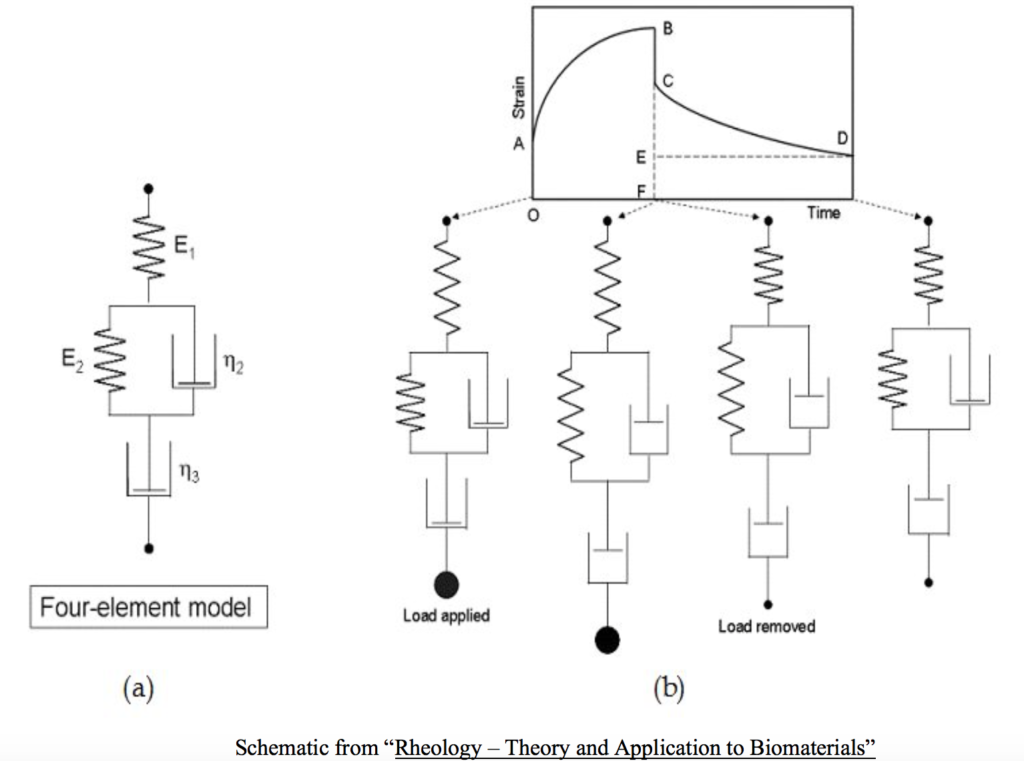
In terms of a muscle’s material continuum the acto-myosin cross-bridge interaction and the behavior it produces is represented by the spring-dashpot system model. The dashpot represents the water component and therefore the force per unit velocity of deformation and the spring represents the force per unit length of deformation. The spring analogy has two components; the series and parallel. The series spring corresponds physically to the passive architecture of the actin and myosin configuration, the sarcomeric Z-disks, and the passive action of the muscle’s tendon. The parallel spring corresponds to the elastic structures that lie passively parallel to the sacromeres, the fasciae, connective sheaths, and various covering like the endomysium, perimysium, and epimysium. Taken together this model of the viscoelastic behavior of whole striated skeletal muscle is called the Four-Element Model.
Understanding the previous definitions and descriptions of the concept of viscoelasticity is important if one is to attempt a transfer the meaning of the neologism (when built from its constituent components) to the behaviors demonstrated by living muscle under varying conditions. Whole in-vivo muscle behavior can be observed and analyzed from both a structural and functional perspective.
Generally, observations of whole in-vivo muscle are categorized as either passive or active. Observations of muscle under passive conditions are defined as the muscle at rest/relaxed, not under any force except the inertia of the body, completely supported by an external object, and no intentional contraction by the individual. Muscle in a passive condition can be assessed via the tactile senses by conducting a palpation with the force acting perpendicularly to the muscle belly through the skin to assess its tonus, volume, shape or material continuity. Additionally, visual assessment can be obtained via MR or ultrasound. Observations of muscle under active conditions is defined as muscle tension created via the cross-bridging of actin and myosin volitionally by an individual, either iso-angularly/statically by co-contracting the muscles acting on a specific joint or joints, or dynamically changing joint angles(s) in opposition to, or creating negative acceleration, some external force. Muscle in an active condition can be assessed via tactile contact along a continuum from the initiation of the contraction from a resting condition all the way through to its volitionally generated maximum tension generating capability at a variety of joint positions under varying ambient and temporal conditions. Observation and assessment of active muscle include but are not limited to; Invasive and Surface Electromyography, Nerve Conduction Velocity, one-repetition maximum tests, isokinetic tests, dynamometers, and endurance tests.
Representing and imagining viscoelasticity as a linear spring and dashpot/damper, it is concluded that in-vivo skeletal muscle has the ability to alter its state and behavior to a shock absorbing quality like a fluid as well as adjusting its length like an elastomer. Muscle’s stress and strain (length and tension) adjustability under dynamic and static conditions (for a given temperature or pressure) manifests through tactility and visually the appearance that its viscosity and elasticity can be rapidly decreased or increased, deforming and flowing in the way non-biological materials do under forces. However, in-vivo muscle is not measured quantitatively by the techniques that measure these properties for non-biological materials. (note: under standard temperature and pressure a non-biological material cannot alter its viscosity or elasticity via the intrinsic manipulation of its material continuum like skeletal muscle can via the cellular response and nervous system communication. The material’s viscosity and elasticity are inherent in its molecular structure and the specific conditions under which a specific stress is applied.)
“While the behavior of classical springs is a useful starting point for understanding the biological response to mechanical stimuli, the reality is far more complicated due to the contribution of two key factors. First, tissues and cells are not passive materials, but rather have tunable mechanical properties governed by dynamic reorganization of their cytoskeletal networks…. Second, while mechanosensation begins with a mechanical response (strain, stress, and/or intracellular forces) cells transduce the mechanical stimuli into a biochemical output, otherwise known as mechanochemical signaling or mechanotransduction.” Cohen, D.M. and Chen, C.S., “Mechanical control of stem cell differentiation” (October 31, 2008), StemBook, ed. The Stem Cell Research Community, StemBook, doi/10.3824/stembook.1.26.1
“There are different sorts of stress (e.g. shear, torsional, etc.) and materials can respond differently for different stresses.” Wikipedia – Rheology
From this perspective it is tentatively concluded that muscle behavior, although intuitively understood to possess both viscous and elastic properties from observed function and sensation, does not possess these qualities as defined. The living cell’s mechanism of influence on the material behavior of muscle is significantly different than that of the inherent molecular structural influence found in non-biological materials.
Muscle structure and function are a summation of active and passive material interactions. The application of non-biological material behaviors to biological ones breaks down under the strictest interpretations of viscosity and elasticity due to this arrangement (muscle as a mixed material continuum). The biggest issue displaying the disparity in applying non-biological passive material elastic behaviors to active biological materials like muscle is demonstrated by the inverse relationship between a force-velocity (f-v) curve developed for muscle acting under forces versus a non-biological material. Non-biological materials have a f-v curve showing that with increasing velocity there is an increase in force while a muscle f-v curve shows that with increasing velocity there is a decrease in force.
“Hill’s equation shows a hyperbolic relation between P and v. The higher the load, the slower the contraction velocity. The higher the velocity, the lower is the tension. This is in direct contrast to the viscoelastic behavior of a passive material, for which higher velocity of deformation calls for higher forces that cause the deformation. Therefore, the active contraction of a muscle has no resemblance to the viscoelasticity of a passive material.” Fung, Y.C., “Biomechanics – Mechanical Properties of Living Tissues”, 1993, Pg. 400
“The living soft tissue of humans and animals are also nonlinearly viscoelastic in the strain range in which they function normally. Fung, Y.C., “A First Course in Continuum Mechanics”, 3rd Edition, 1994, Pg. 189
It is therefore concluded that in-vivo muscle is a non-linear, pseudo-viscoelastic material. This means that although visual observation and tactile sensory feedback of muscle behavior under forces may give the resemblance and feel of the viscous ‘fluid-like’ and elastic ‘spring-like’ behaviors that are observed in passive non-biological materials, this is an imitation of these properties. It is very difficult to establish highly predictable and proportional changes in each variable as conditions change for the muscle both internally and externally. Practitioners working with muscle in the field are not taking quantitative measurement of these properties and are only observing the summation of the living cell activity. The active and passive components are concomitantly linked, and their behaviors coupled making it impossible to know which component is contributing more or less to normal and abnormal movement due to the complexity of this relationship. It is deemed therefore that the term “viscoelasticity” is applied as a psycholinguistic pragmatic in order to try and make sense of the various observations of in-vivo muscle behaviors on the human lever system. The property of viscoelasticity does not accurately represent the true and full description, or understanding, of actual living muscle behaviors.
“The exact contribution of each of the anatomic elements to muscle stiffness, or the resistance to deformation, is unknown. The contribution of the contractile elements appears to be related to the velocity of the deformation. As muscle is deformed or stretched, the contribution from the non-contractile elements increases.” Magee, Zachazewski, Quillen – Scientific Foundations and Principles of Practice in Musculoskeletal Rehabilitation 2007, Pg. 100
When considering the effect that forces have on the deformation of materials the property of stiffness is discussed along with the property of elasticity.
“In studies of motor control and coordination, stiffness is often considered the most relevant property of the movement actuators.” Ettma, G., “Modeling Skeletal Muscle-Tendon Units as Actuators in Motor Behaviors: Morphological Aspects”, Corpus, Psyche et Societas 5, 22-48 1998
Stiffness can be thought of in both its noun and adjective forms:
Stiff (adj.) – not easily bent or changed in shape; rigid.
Stiffness (n.) – the inherent physical property of a thing that does not easily bend.
“Stiffness is the resistance of an elastic body to deformation by an applied force along a given degree of freedom (DOF) when a set of loading points and boundary conditions are prescribed on the elastic body. Generally speaking, the elasticity of a material is not the same as that material’s stiffness. Elasticity is a property of the constituent material; stiffness is a property of a structure. The elasticity is an intensive property (its material construction without consideration of size, volume, shape) of the material; stiffness is an extensive property of the solid body dependent on the material, its shape and the boundary conditions.” Wikipedia
It is important to distinguish the adjective ‘stiff’ from the noun ‘stiffness’. Applied to muscle the term ‘stiffness’ defined above as an extensive property would refer to its cross section, shape and length. The professional literature and lay descriptions of lever system movement can cause confusion regarding to which usage – noun or adjective – the term ‘stiffness’ is applied. Whether an external force is acting directly on the mass of muscle thus deforming it in the direction of the force – i.e. by a tactile palpation of a muscle belly or by a foam roller – or an external force acting on a lever system of which the muscle is an active component producing torque – i.e. bending the elbow with a dumbbell in one’s hand – the two represent entirely different descriptions of state conditions to which the term stiffness can be applied. As a vernacular term ‘stiff’ might be a subjective description that simply means the lever system doesn’t want to move as easily as one desires. This does not necessarily imply that stiffness and tightness are synonymous. Whether muscle does or does not want to change its state depends on the internal friction of its various fluids (flow), the molecular structure of its various connective tissue (length/stretch), the nervous system’s influence (tension), and the architecture of the various active elements, none of which can be subject independently to the strictest application of the terms defining viscoelasticity.
As initially stated the context of this document falls under the muscle system’s active contribution to efficient movement or maintenance of position occurring within the human lever system acting under forces within an exercise paradigm. The passive elements: titin, collagen, fascia, etc. and what is currently known regarding their contribution to the structure and function of skeletal muscle are acknowledged and appreciated. However, it is unclear that they are equal partners in producing and managing efficient movement or maintenance of position. An argument can be made that in order for the passive tissue properties (their storage and expression of elastic energy as a mechanical output) to contribute to the efficiency of movement, the active contractile elements must be functioning optimally in order to create and transfer any elastic energy stored in them to the tendon. (The tendon represents the final output point of the system producing torque on the lever system.
This is readily apparent in neurological damage that results in diminished motor function (i.e. stroke, peripheral nerve damage). This kind of deficit displays the reality that stored and expressed elastic energy in passive tissues alone cannot initiate or sustain efficient movement. Active muscle contraction provides the internal anchoring – via the tension it produces – to the passive tissue scaffolding. This active tension establishes the foundation for the storage and expression of passive tissue’s elastic energy and its contribution to movement control, joint mobility, and joint stability. This possibly places the active elements as significantly more influential and important to motor control and movement in any hierarchy of tissue importance. Any sensorimotor process that supports the development and sustenance of active muscle tension is of critical import.
“A third type of control strategy is related to both feedback and feedforward control and involves modulation of the ‘tone’ in specific muscles to provide an underlying degree of stability to the joints. This activity increases the stiffness of muscles that surround the joints.” Richardson, Hodges, Hides, “Therapeutic Exercise for Lumbopelvic Stabilization – A Motor Control Approach for the Treatment and Prevention of Low Back Pain.” 2nd Edition, Page 25.
“Johansson et al (1991a,b) have undertaken much of the neurophysiological research linking muscle stiffness to joint stability. They have described muscle stiffness as having two components: intrinsic and reflex mediated stiffness.” Richardson, Hodges, Hides, “Therapeutic Exercise for Lumbopelvic Stabilization – A Motor Control Approach for the Treatment and Prevention of Low Back Pain.” 2nd Edition, Page 78.
“Reflex Mediated Stiffness depends on the excitability of the motor neuronal pool, which in turn is dependent on the primary spindle afferents set by the degree of stretch of the muscle and the activity of the fusimotor neurons.” Richardson, Hodges, Hides, “Therapeutic Exercise for Lumbopelvic Stabilization – A Motor Control Approach for the Treatment and Prevention of Low Back Pain.” 2nd Edition, Page 78.
“The results indicate that both hamstrings and quadriceps demonstrate a significant increase (greater than 100%) in their antagonist coactivation pattern during the final 40 degrees of fast extension and flexion movements, respectively, as limb velocity increases. A minor decrease in antagonist activity of the hamstrings (24%) and quadriceps (8%) was evident during the initial phase of the extension and flexion movements, respectively, as joint velocity increased. We concluded that as limb velocity is increased, there is a substantial reflexive (unintentional) increase in the contribution of the antagonist musculature to joint stiffness and reduction of laxity”. Hagood, Solomonov, et. al, 1990
In conclusion, it is apparent that empirical observation of active and passive muscle function, extrapolated from observing lever system (limb and trunk) motions under forces, gives the appearance of muscle expressing the viscous, elastic and stiffness properties found in non-biological materials subject to external and internal forces. If anything, it seems as though the literature uses the terms viscosity, elasticity and stiffness in their noun forms applied to the entire lever and muscle system behavior considering them as one continuous material entity, and sometimes in their adjective forms to qualitatively describe an attribute of movement.
The unique structural makeup of muscle with its integrated active and passive tissue capabilities, under the strong influence of the central nervous system, makes it difficult to completely separate each property for analysis under in-vivo active or passive conditions. This makes it difficult to measure each property and therefore reach a definitive conclusion about the change in a given property contributing to an observed dysfunction or restoration of motor control and/or motion deficit.
“The knowledge of the mechanical pathway, through which the body level loads are distributed between organs, then within the tissues and further along the extracellular matrix and the cells, is critical for the success of various interventions. However, this information is not established.” Erdemir, A., “Predicting Cell Deformation from Body Level Mechanical Loads”
However, given (1) that the active component (tension from the muscles cells transferred to the tendon) contributes to movement and its influence on change within its normal physiological operating window, (2) that muscle cell tension provides the internal stability from which passive tissues can store and transfer their elastic energy, and (3) that the expression of the two are regulated by the central nervous system via the motor unit, it may be that assessing the contractile apparatus’ ability to generate torque via the lever system is the simplest window into assessing the effectiveness of movement system performance. This supersedes attempts to evaluate the in-vivo passive tissue elasticity since this can only be measured at the most extreme ends of the tissue system length and even then, due to reflex mediated factors acting on the contractile components, it is impossible to remove the muscle contractile contribution to, and conclusions about, the muscle exhibiting normal or abnormal properties of: total stiffness, elasticity or viscosity – let alone gauge any sense of change in these properties solely from changes attributed to passive tissues.
A distinction needs to be made regarding the changes in the quantitative elasticity of tissues brought about by external forces acting to change their length from a molecular structure “non-biological fabric” paradigm (compress/shorten or stretch/lengthen) and changes in tissues due to the internal cellular adaptations driven by RNA expression. Wolff’s and Davis’s Laws reflect the position that the cells of bones and soft tissues adapt IAW with the direction, intensity and duration of the force acting chronically on the tissues through the cellular machinery that alters the morphology of the cell and results in any change in tissue length, density or volume from the inside out.
“Tendon fibroblasts expressed higher levels of a-smooth muscle actin when they were oriented parallel to the stretching direction than when they were oriented normal to the stretching direction. Also, cyclic stretching of the fibroblasts perpendicular to their orientation induced a higher activity level of secretory phospholipase A2 compared with stretching of the cells parallel to their orientation. Thus, these results show that fibroblast responses to mechanical stretching depend on cell orientation to the stretching direction.” James H.-C. Wang, Guoguang Yang, Zhaozhu Li, Wei Shen, “Fibroblast responses to cyclic mechanical stretching depend on cell orientation to the stretching direction”, Journal of Biomechanics 37 (2004) 573–576
Cells hypertrophy, atrophy, proliferate, or through apoptosis – die. This can be considered a form of plasticity but not the type of plasticity attributed to non-biological materials when forces exceed their yield stress limit. As an example, a type of plasticity exhibited by in-vivo tissues is reflected in the changes to nervous system function. Changes in neuron-to-neuron communication are altered by modifications in dendritic, synaptic or axonal properties, which are derived from internal cell messaging.
“Plasticity, for example, is used to refer to changes either at the cellular/network level (neural plasticity) or at the behavioral level (behavioral plasticity). The underlying mechanisms of neural plasticity are multiple; they include biochemical, structural, and functional changes.” Freund, Sabel, Witte, “Advances in Neurology, Volume 73 – Brain Plasticity”, 1997, page 297
Another example is shown in the increase of sarcoplasmic reticulum volume that contribute to muscle cell hypertrophy following several bouts of resistance training over several weeks. Cellular structural integrity is maintained during the progressive biological plasticity process transferring a benefit to the material whereas molecular structural integrity is lost in non-biological material plasticity leading to a permanent deficit in its capability.
“Chronic loading in the form of physical training leads both to increased collagen turnover as well as, dependent on the type of collagen in question, some degree of net collagen synthesis.” Kjaer, M., “Role of Extracellular Matrix in Adaptation of Tendon and Skeletal Muscle to Mechanical Loading”, doi: 10.1152/physrev.00031.2003, Physiol Rev April 2004 vol. 84 no. 2 649-698
“This experiment extends knowledge of human tendon apoptosis by showing that apoptosis can occur in response to short term, high strain mechanical loading. This is the first report of mechanical loading of intact tendon causing excessive apoptosis.” A Scott, K M Khan, J Heer, J L Cook, O Lian, V Duronio, “High strain mechanical loading rapidly induces tendon apoptosis: an ex vivo rat tibialis anterior model”, Br J Sports Med 2005;39:e25 doi:10.1136/bjsm.2004.015164
“Responses to mechanical stimulation involve multiple and redundant pathways 2,3. Transduction of mechanical load to a biochemical signal involves Ca2+ signaling (Ca2+ influx, efflux, and release from intracellular stores), IP3 and perhaps Ca2+ diffusion through gap junctions, purinoceptor activation, and mechanosensitive ion channels. These immediate signaling events to load may be useful in regulating the down-stream effects of mechanical stimulation on connective tissues. Some positive outcomes of loading are increased matrix gene expression 93,94, mitogenesis 95,96, bone mineralization97, and blocking cytokine actions98. Some negative effects of load are apoptosis99, loss of membrane integrity100, and upregulation of cytokines and matrix metalloproteinases 58,63. Lack of loading altogether can lead to bone resorption and disuse osteoporosis 97,101. However, the cell responses to load are dose-sensitive, cell-type sensitive, and synergistic or antagonistic with pathway activation/ inactivation.” Wall, M.E., Banes, A.J., “Early responses to mechanical load in tendon: Role for calcium signaling, gap junctions and intercellular communication”, J Musculoskelet Neuronal Interact 2005; 5(1):70-84
From the molecular structure based “non-biological fabric” paradigm, changes in elasticity due to an acute bout of external force application attempting to alter lever orientations by changing passive and active tissue length with the intention of changing the integrity of the tissue continuum, independent of the internal cellular mechanism driving tissue morphological change, may actually be creating plastic deformations to tissues due to damage of the cell-to-cell integrity. This may compromise its innate non-linear pseudo-viscoelasticity properties and the precise development and expression of stored mechanical energy that is the hallmark of efficient human movement. A tissue’s current viscosity, elasticity and stiffness – if these terms are allowed to be loosely applied to in-vivo human muscle – is actually a function of the internal cellular driven changes in morphology over time. Healthy tissue length change occurs because cells grow, shrink or die – not because cells are stretched and stay stretched or compressed and stay compressed, as in non-biological materials plastically deformed. This is not the same as a lever system’s resistance to, or acceptance of, the moment-to-moment length change as a function of the nervous system’s control of tension via the active elements delivered to the tendon. This is where muscle behavior takes on the appearance and feel of a non-linear “spring-like” quality, keeping in mind that the process leading to the development of reaction force in a non-biological spring is not the same as the reaction force generated in a muscle. Ultimately it is the muscle’s active element’s change in tension that changes lever system orientations, or maintains positions, within the system’s current physiological operating window.