News & Articles
A General Description of the Central and Peripheral Nervous Systems
The CNS is made up of a diverse array of interconnected neurons (among other things, i.e. astrocytes, glial cells) located in the brain and spinal cord. The CNS generates and dispatches motor control signals (This is termed “Efferent” information) and receives and processes sensory information from the peripheral nervous system (PNS) (This is termed “Afferent” information) and autonomic nervous system (ANS) in order to exercise control of the body.
“The CNS is described as having “seven major divisions”; Kandel, Schwartz, Jessel – “Principles of Neural Science”, 4th Edition, Page 317
Listed from Most Cephalad to Caudal Structures)
- The Two Cerebral Hemispheres
- The Diencephalon
- The Cerebellum
- The Mid Brain
- The Pons
- The Spinal Cord
The CNS is a principled system constituted by an organization built from neuronal interconnectivity. The seven major divisions of the CNS listed previously are the key hubs of a functional network with specified hierarchical relationships. These neuron-to-neuron connections via their dendritic and axonal projections create configurational pathways that link together the individual components of the system. The specific locations and the orderly manner in which these pathways connect the various regions of the CNS, PNS, and ANS creates a topographical map of the system – within the system – called somatotopy. This somatotopic map involves the different divisions (and their sub-components) in a unique manner so that different types and rates of information flowing within and throughout the system can be processed and managed with precision and thus create meaning.
“Somatotopy is a feature of receptive field arrangement. The body is represented in the nervous system in an orderly arrangement which preserves the relationships of the body parts. The rule is that adjacent body parts are represented in adjacent sites of the nervous system.” Butler, David, S., “The Sensitive Nervous System”, 2001, Page 30
“Examination of the major sensory or motor pathways reveals a highly and tightly organized nervous system. In particular, at each of many levels, there are fairly exact maps of the world within the brain. For example, sensory axons within its ascending pathway are arranged in a very orderly manner, with fibers from specific anatomic locations (e.g., digits, hand, forearm, etc.) preserving very specific topographical locations within the dorsal column, thalamus, internal capsule, and sensory cortex.” JOURNAL OF SURGICAL ORTHOPAEDIC ADVANCE VOLUME 17, NUMBER 3, FALL 2008 1161
The functional systems and their relationship to the body are such that most neural pathways share a bilaterally symmetry (similarity). Each of the cerebral hemispheres primarily controls the contralateral body structures – via decussation (i.e. The right cerebral hemisphere generally controls the muscles of the left side of the body and the left hemisphere generally controls the right side).
“Decussation describes the point where the nerves cross from one side of the brain to the other, and typically the nerves from the left side of the body decussate to the right side of the brain and the nerves from the right side of the body decussate to the left brain, however depending on the function of the nerves the level of decussation is variable.” Wikipedia
Corpus Callosum
The Corpus Callosum is prominent commissural network of neural fibers that structurally and informationally connects the symmetrical regions of the left and right cerebral hemispheres together. An important feature of this structure is the connection it makes between the somatosensory cortical areas of the right and left cerebral hemispheres. This creates the network information construct so that the CNS can process sensory information in a way that builds a complete 4-dimensional map of the living system thus allowing it to leverage the control inherent in the physics of CNS bilateralism. The system knows where, how, and when it is.
“Thus, one of the functions of the corpus callosum and the anterior commissure is to make information stored in the cortex of one hemisphere available to corresponding cortical areas of the opposite hemisphere.” Guyton, A. C. “Textbook of Medical Physiology”, 9th Edition, 1996, Page 742
“These motor zones are endowed with dense callosal connections (Ebner and Myers 1965; Jones and Powell 1968; Pandya and Vignolo 1969; Karol and Pandya 1971; Jones et al. 1979; Pappas and Strick 1981; see also Gould et al.1986) which may enable homotopic zones related to axial movements to function as a co-ordinated unit like their sensory counterparts. This possibility is suggested by the finding that direct microstimulation of callosal fibres reciprocally connecting the motor areas of the two hemispheres produces symmetrical and bilateral contractions of axial muscles (Spidalieri and Guandalini 1983). The unitary representation of body midlines ensured in the bi-hemisphered brain by the corpus callosum could therefore have a fundamental role in the bilateral activation of axial muscles and participates, together with other sensory cues, in the central mechanism constituting the reference axis to direct the body and mobile appendages in the external space.” T. Manzoni, et.al. “Experimental Brain Research”, 1989, 76:251-266
“An interesting finding in some chronic musculoskeletal pathology is bilateral dysfunction in unilateral injury (Bullock-Saxton, Janda, and Bullock 1994; Cools et al. 2003; Roe et al 2000; Wadsworth and Bullock-Saxton 1997; Wojtys and Hutson 1994). Bullock-Saxton, Janda and Bullock (1994) found that subjects with chronic ankle sprain exhibit altered muscle activation patterns on both the injured and uninjured sides.” Page, P., Frank, C., Lardner, R., “Assessment and Treatment of Muscular imbalance: The Janda Approach”. Page 25, 2010
Cerebral Cortex
The Cerebral Cortex is the relatively thin and most outer layer of each of the cerebral hemispheres and is posited to be responsible for the planning and execution of human action. The Cerebral Cortex is divided anatomically into four lobes (listed anterior to posterior); frontal, parietal, temporal and occipital. This lobed and layered is organized in such a way as to provide efficient input-output communication between the various cortical neurons. The neurons that constitute each lobe and layer project to different parts of the brain and spinal cord. It has been determined that this structure fundamentally processes the afferent sensory information and initiates and sends efferent motor commands.
The concept of it being a primary input/output processor gives it the distinction of being … “the seat of consciousness.” “Somatosenory Processing – “From Single Neuron to Brain Imaging”. Rowe, M. J., Iwamura, Y., 2001, Page 131)
“Indeed, it is impossible to understand anything about how the brain works without conceding that its main problem is to put mass into motion. Instead of saying ‘mass’ one might say ‘inertial moment’, or the considerable complex forces that arise as soon as a mass is in motion… The Russian physiologist Bernstein drew attention to the fact that in animals and in man, the limbs have enormous ‘degrees of freedom,’ and consequently nature has had to find tricks to simplify the work of the brain. Study of the geometry of movements helps to grasp the natural solutions devised by the nervous system to solve this problem. Skeletal anatomy can be explained in this context; moreover, pre-wiring motor synergies is another way of simplifying neuro-computation, and simple kinematic relationships connect the geometry of movement and dynamics” (Berthoz, 1997)
General Description of the Peripheral Nervous System (PNS)
The PNS is demarcated from the CNS by defining itself as that part of the nervous system that contains all the neurons that lie outside of the CNS. Although anatomically distinct it is not functionally separate from the CNS. The peripheral nervous system (PNS) connects the central nervous system (CNS) to sensory organs (such as the eye and ear) and other organs of the body; muscles, joints, blood vessels, skin and glands.
The peripheral nervous system includes the 12 cranial nerves, the spinal nerves and their roots, and the autonomic nerves that are concerned with the regulation of the organs, i.e. the pancreas, the liver, the heart, the smooth muscle in blood vessel walls, and the glands.
Biological Tissue Continuum
The human body and the total of all of its tissues is a material continuum. A material continuum is described as a continuous, non-homologous distribution of mass with a distinct boundary that fills a definable space. The body constitutes a diverse array of cells and tissues each with its own degree of consistency as measured by the quantity of mass per unit volume for each area of unique tissue expression (i.e. lungs, liver, bone, skin). Therefore, as a material continuum, the human body is subject to analysis by the field of physics/mechanics. The muscular and nervous tissue systems are both components of, and integrated materials within, the fabric of the body’s total material continuum.
“At the cellular, tissue, organ, and organism level it is usually sufficient to take Newton’s laws of motion as an axiom.” Fung, Y.C. “Biomechanics: Mechanical Properties of Living Tissues”, 2nd Edition, 1993, Page 23
Sensory Receptors
Sensory Receptors are a diverse array of specialized nerve cells with terminal endings, referred to as boutons, embedded within the body’s material continuum. These specialized neurons change their polarity when subjected to a mechanical, thermal, chemical, and/or electrical stimulus. If the stimulus is strong enough then a threshold is reached and results in depolarization of the cell which generates and sends an action potential into the CNS informing the system of the presence of the stimulus. The signal is termed “Afferent”.
“Sensory receptor cells have many common characteristics. All have the same task: to convert a stimulus into an electrical signal in a form that can be used to alter behavior.” Fain, Gordon L., “Sensory Transduction”, 2003, Page 19
This information can also be generated indirectly by second-order neurons that propagate a signal to the CNS for interpretation. Second order neurons carry the information from the central nervous system to the thalamus. Information traveling along second order neurons deccussates from one side of the CNS to the other side of the CNS.
“Two important conclusions about signaling in the nervous system are (1) that the nerve cells act as building blocks for perception and (2) that the abstract significance of the message can be extremely complex, depending on the inputs a neuron receives.” Nicolls, Martin, Wallace, Fuchs,“From Neuron to Brain”, 4th Edition, 2000, Pg 19.
Receptor Types
Sensory neuron receptor somas are located in the spinal cord dorsal root ganglion. Sensory receptors are often named by their discovery (i.e. Golgi organs, Pascinian Corpuscles) classified according to the type of energy stimulus. The stimulus creates a transformational change in a neuron’s bouton which may depolarize the cell and report a signal to the CNS. Sensory Receptors generally fall under two broad classifications:
Exteroreceptors – transduce information from influences exogenous to the body’s material continuum. (glabrous/cutaneous)
Interoreceptors – transduce information from influences endogenous to the body’s material continuum. (visceral/organ)
Within these two broad categories sensory receptors can be further classified according to the primary energy stimulus acting on the bouton for which each is attuned. (although some receptors are poly-modal meaning that they depolarize to a variety of stimulus types).
- Thermoreceptors – Thermal Energy
- Electroreceptors – Electrical Energy
- Mechanoreceptors – Mechanical Energy
- Baroreceptors – Pressure Energy
- Chemoreceptors – Chemical Energy
- Nociceptors – Noxious Stimulus (from diverse energy influences)
- Magnetoreceptors – Magnetic Energy
- Photoreceptors – Photon Energy
- Hydroreceptors – Ambient Humidity
- Osmoreceptors – Chemical Concentrations in Fluid
- The receptor types directly* associated within the muscular tissue are:
- Free Nerve Endings
- Paciniform Corpuscles
- Golgi Tendon Organs
- Muscle Spindles (Intrasfusal Fibers)
- Meissner’s Corpuscle
- Merkel Cells
- Ruffini’s Corpuscles.
* – The term “directly” here infers that the afferent signal generated by the receptor synapses without interruption or modification by an internuncial/interneuronal cell between it and the alpha motor neuron.
“… , each Ia afferent fiber makes contact with over 90% of homonymous motoneurons.” Macintosh, Gardiner, McComas, “Skeletal Muscle: Form and Function”, 2006, Page 47.
“Moreover, it appears that the switching interneurons operate (fire) under control of various proprioceptive and descending control signals and hence significantly contribute to the shaping of the locomotor pattern (timing phase transitions, shaping motoneuronal firing bursts, etc.)” Ivashko, et al. Modeling Neural Control of Locomotion: Integration of Reflex Circuits with CPG, ICANN 2002 LNCS 2415, pp.99-104
These sensory receptor types have the capacity to impact the alpha motor units of the muscle tissue within which they are embedded. The Muscle Spindle plays an integral role in motor system control in that it communicates changes in a muscle’s length and velocity directly back to alpha motor neurons in the Ventral Horn of the spinal cord portion (known as a Ia afferent) of the CNS and also to the brain via interneurons and ascending spinal tracts.
“The primary function of proprioceptive reflexes in regulating voluntary movements is to adjust the motor output according to the biomechanical state of the body and limbs. This ensures coordinated pattern of motor activity during an evolving movement, and it provides a mechanism for compensating for the intrinsic variability of motor output.” Kandel, Schwartz, Jessel – Principles of Neural Science, 2000, Page 727
“These connections provide a means for rapidly executed reflex adjustments in muscle length following any stretching or change in muscle load.” Macintosh, Gardiner, McComas, Skeletal Muscle: Form and Function”, 2006, Page 47
A muscle spindle’s intrafusal fibers are wholly unique amongst the other receptor types in that its sensitivity (threshold setting to depolarize) is influenced by efferent information generated by the CNS. The CNS subsystem that influences the sensitivity is called the Fusimotor System. A spindle’s afferent information directly acts on specific alpha motoneurons to adjust their firing frequency and thus total tension development (Called homonymous alpha motor neurons). All the other sensory receptor sensitivity thresholds for a given stimulus are not known to be directly influenced and adjustable by the CNS.

Description of Fusimotor System (muscle spindle intrafusal fibers)
The fusimotor system innervates the intrafusal fibers of a given muscle spindle by efferent motor neurons located in the ventral horn of the spinal cord. These are termed gamma motor neurons.
“Three reasons have been adduced at various times to explain the existence of a specific fusimotor system; these shade into each other and can still all be accepted in some degree, although in their extreme forms they are exclusive. First, fusimotor activity may provide for the maintenance of the sensitivity of the spindle in the face of externally imposed changes, most notably prolonged change of muscle length or phasic effects of muscle contraction acting to unload the spindle. The former may well apply to the sensitizing action of the dynamic axons in maintaining a high sensitivity to small stimuli. Second, fusimotor activity seems to provide for the regulation of the sensitivity of the spindle to suit the prevailing mechanical conditions, rather than simply holding it at some universally appropriate value. This may be dubbed parameter control, leading to the question of which parameters of spindle responsiveness should be viewed as the ones under control. As just noted, on the basis of sinusoidal stretching it no longer seems appropriate to believe that the dynamic axons exert a specific action on the velocity sensitivity of the primary ending independently of its position sensitivity, because when measured under dynamic conditions the latter also is raised by dynamic action. Rather, the gain of the ending seems to be under control, being high during dynamic action and low during static action irrespective of the size of the stimulus under consideration. Third, fusimotor activity biases the spindle and increases the firing rate of both primary and secondary endings for a given length of the muscle.” Matthews, P. B. C. 2011. “Muscle Spindles: Their Messages and Their Fusimotor Supply”. Comprehensive Physiology. 189–228.
Alpha-Gamma Co-Activation
The dynamic connectivity and resulting interdependent information flow of the sensory and motor system interface is nowhere as plain to be observed then in the process of alpha-gamma co-activation. The partnered actions between alpha motor neurons and gamma motor neurons is tightly linked in that both alpha and gamma motor neurons are influenced simultaneously.
“… in voluntary movements, the reference value is set by descending signals that act on both the alpha and gamma motor neurons.” Kandel, Schwartz, Jessel – “Principles of Neural Science”, 4th Edition, Pg. 728
This interdependency ensures that alterations in the tension development and any changes in length of the extrafusal fiber is governed by the ability to monitor and tune that output via intrafusal fiber Ia feedback to the CNS.
“… the alpha motor neurons activate the extrafusal fibers to produce the force required for the task, and the gamma motor neurons activate the intrafusal fibers to set the desired level of feedback from the muscle spindles.” Enoka, Roger M., Neuromechanics of Human Movement, 4th Edition, 2008, Pg. 252
The healthy behavior of the motor control system via muscle tension development and its response to dynamic and static conditions is contingent on the integrity of the interdependent nature of the sensory-motor system relationship.
“In the absence of sensory feedback, however, the maintenance of fine control tends to deteriorate as the task progresses, possibly because of the accumulation of small errors.” Nicholls, John G., et.al., “From Neuron to Brain” , 4th Edition 2001, Pg. 459
“This means that the level of activity of a spindle receptor depends not only upon the actual muscle length (and velocity, for primary receptors) but also upon the level of muscle activation.” Latash, Mark L., “Neurophysiological Basis of Movement”, 1998, Pg. 195
Information Flow and the Sensory-Motor Control System
The central motor system and the peripheral sensory apparatus are intimately
linked together in the spinal cord and brain via the network of neuron soma projections (axons and dendrites) and their respective synaptic connections. The constant ebb and flow of information within and between these two systems contribute substantially to the development of muscle tension and coordination of motor behaviors under a variety conditions.
There are four basic classifications of information distribution and governance that describe the information exchange throughout and between the systems: Excitation, Inhibition, Convergence, and Divergence.
Excitation (n.)
The activity produced in an organ, tissue, or part, such as a nerve cell, as a
result of stimulation. In a neuron it represents the change in its polarity that produces a receptor or action potential.
Inhibition (intr. v.)
The blocking or limiting of the activity of an organ, tissue, or cell of the body, caused by the action of a nerve or neuron or by the release of a substance such as a hormone or neurotransmitter. In a neuron, it represents the change in its polarity that restricts the development of a receptor or action potential.
“The role of inhibition in the working of the central nervous system has proved to be more and more extensive and more and more fundamental as experiment has advanced in examining it. Reflex inhibition can no longer be regarded merely as a factor specially developed for dealing with the antagonism of opponent muscles acting at various hinge-joints. Its role as a coordinative factor comprises that, and goes beyond that. In the working of the central nervous machinery inhibition seems as ubiquitous and as frequent as is excitation itself. The whole quantitative grading of the operations of the spinal cord and brain appears to rest upon mutual interaction between the two central processes ‘excitation’ and ‘inhibition’, the one no less important than the other. For example, no operation can be more important as a basis of coordination for a motor act than adjustment of the quantity of contraction, e.g. of the number of motor units employed and the intensity of their individual tetanic activity. This now appears as the outcome of nice co-adjustment of excitation and inhibition upon each of all the individual units which cooperate in the act.” Sir Charles Scott Sherrington, “Inhibition as a Coordinative Factor”, Nobel Lecture (12 Dec 1932). Nobel Lectures: Physiology or Medicine 1922-1941 (1965), 288.
Convergence (n.)
A postsynaptic nerve cell or pool of cells – called the target – receive the sum of input information originating from one, or several, exogenous presynaptic nerve cells. This process alters and reduces the large information flow towards a single smaller common point for a particular result.
“… because there are usually five to 10 times more sensory neurons than motor neurons, a single motor cell typically receives input from many sensory cells. This pattern of connection, called convergence, is common at the output stages of the nervous system.” Kandel, Schwartz, Jessel – “Principles of Neural Science”, 4th Edition, Pg. 26
Divergence (n.)
When a specific neuron or neuron pool depolarizes and distributes chemical and electrical information via its dendrites and axons to the surrounding network of neurons which then widens the distribution of this initial generation of information this is referred to as Divergence.
“This pattern of connection, in which one neuron activates many target cells, is called neuronal divergence; it is especially common in the input stages of the nervous system.” Kandel, Schwartz, Jessel, “Principles of Neural Science”, 4th Edition, Pg. 26
The living nervous system is constantly sending, receiving, processing, revising,
and editing chemical and electrical information within the body’s material continuum. These four fundamental processes describe the basic mechanisms by which sensorimotor information is created, integrated, and managed to control muscle function. See Sherrington’s Final Common Path Conclusion.
“All neurons within the central nervous system take account of influences arriving from diverse inputs to create their own new messages with new meanings. The term integration was introduced by Sherrington.” Nicholls, John G., et.al., “From Neuron to Brain”, 4th Edition 2001, Pg. 18
The total information set represented at any point in time from the peripheral sensory receptor system into the CNS provides, among other neural processing inputs, reflects the 4- dimensional account (the 3 cardinal planes of space and the time dimension) of the body’s configuration to itself as well as within it’s external environment.
“The nervous system is unique among human body systems in its spatial and temporal organization. The central nervous system is divided into highly specialized regions that have unique properties in terms of molecules, cell types, connections, and functional systems. The functions of these units vary with time, spanning the gamut from the millennia of evolution to the millisecond choreography of neurophysiologic events. This temporal and spatial specialization is well suited to the application of informatics techniques. In fact, such methods will be required as the basis for beginning to understand and organize the ever-increasing amount of neuroscientific information that is accumulating about this, the most complicated system known. What is ultimately required is a multidimensional database organized with three dimensions in space and one in time along with a seemingly infinite number of attributes referable to these four physical dimensions.” JOHN MAZZIOTTA, MD, PHD, et al., A Four-Dimensional Probabilistic Atlas of the Human Brain, J Am Med Inform Assoc. 2001 Sep-Oct; 8(5): 401–430. PMCID: PMC131040,
“In modern physics, space and time are unified in a four–dimensional Minkowski continuum called spacetime, whose metric treats the time dimension differently from the three spatial dimensions (see below for the definition of the Minkowski metric/pairing). Spacetime is not a Euclidean space.” Wikipedia
The Motor Response and Action Continuum – How a Stimulus Elicits a Motor Response via Reflex Arcs and Automatic Processes.
Reflex (n.) – an action that is performed as a response to a stimulus and without conscious thought.
“The unit reaction in nervous integration is the reflex, because every reflex is an integrative reaction and no nervous action short of a reflex is a complete act of integration.” Charles S. Sherrington – “The Integrative Action of the Nervous System” 1906, Pg. 7
Arc (n) – a curved shape, or something shaped like a curve.
“Reflexes are coordinated, involuntary motor responses initiated by a stimulus applied to peripheral receptors.” Kandel, Schwartz, Jessel – “Principles of Neural Science”, 4th Edition, Pg. 735
“The noun ‘reflex’ means something that reflects or is a consequence of something … a reflex is a muscle contraction induced by an external stimulus that cannot be changed by “pure thinking’, that is, by a volitional act that is not accompanied by another muscle contraction.” Latash, Mark L., “Neurophysiological Basis of Movement”, 1998, Pg. 64
“The afferent feedback from sensory receptors to the CNS can evoke rapid responses that are known as reflexes (Pierrot,-Deseilligny & Burke, 2005).” Enoka, Roger M., Neuromechanics of Human Movement, 4th Edition, 2008, Pg. 257
Reflex Arc (n.) – the nerve pathway involved in a reflex action including at its simplest a sensory nerve and a motor nerve with a synapse between.
A simple reflex arc is a system of specifically interconnected neurons consisting minimally of an afferent nerve with its sensory receptor, that when stimulated acts on a nerve center (central processor – interneuron or association neuron), which in turn acts on an efferent nerve, called the effector, that stimulates the associated muscle or gland to some action that occurs in repeatable and predictable way when the same stimulus is applied to the receptor. This is often called a monosynaptic reflex arc.
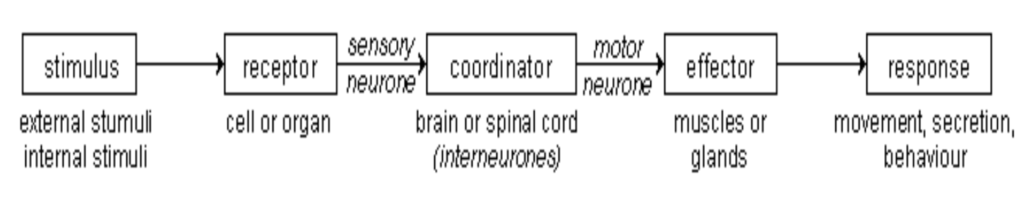
“For our purpose the receptor is best included as a part of the nervous system, and so it is convenient to speak of the whole chain of structures – receptor, conductor, and effector – as a reflex arc.” Charles S. Sherrington – “The Integrative Action of the Nervous System” 1906, Pg. 7
Without reflex arc systems built into the CNS governance of motor control many motor behaviors would not be possible, or would degrade markedly over time, especially those requiring accurate and precise actions. Reflexes are robust and necessary building blocks for orchestrated human movement and position maintenance.
“Co-ordination, therefore, is in part the compounding of reflexes.” Charles S. Sherrington – “The Integrative Action of the Nervous System” 1906, Pg. 7
“These reactions, in which there follows on an initiating reaction an end-effect reached through the mediation of a conductor, itself incapable of either of the end-effect or, under natural conditions, of the inception of the reaction, are ‘reflexes.’” Charles S. Sherrington – “The Integrative Action of the Nervous System” 1906, Pg. 6
For many years following the discovery of reflex systems it was concluded that motor behaviors facilitated with reflex arcs could not be altered or modified from their intrinsic contributions to the overall governance of motor behavior under normal conditions. Recent research in this area has demonstrated this assumption to be untrue.
“It now is clear that even so-called reflex arcs lie within modifiable microcircuits (Marder, 2011), as already forecast in the work of Sherrington and embedded in the ideas of William James.” “Neural Circuits Underlying Emotion and Motivation: Insights from Optogenetics and Pharmacogenetics”, Edited by Anton Llango and Mary Kay Lobo, February 2014, Volume 8, Article 39, Pg. 126.
“We now know that under normal conditions reflexes can be modified to adapt to the task.” Kandel, Schwartz, Jessel – “Principles of Neural Science”, 4th Edition, Pg. 714
Reflex circuitry is classified by the amount of connections with, and influences over, other neurons. I.e. monosynaptic, oligosynaptic, and polysynaptic. Their anatomical location in the nervous system also contributes to classification. I.e. spinal cord or brain. The neuronal configurations that construct the many varieties of reflex arcs where skeletal muscle is the effector organ tend to be located in the spinal cord and are called “spinal reflex arcs”. The preceding descriptions are based on the findings of the artificial and highly controlled laboratory environment under conditions that likely do not readily represent normal in-vivo functionality under diverse environmental situations. As is reflected in the quote below – regarding research in this area – the application of a low-intensity magnetic stimulus to the CNS in order to alter regional electrical conditions and ultimately alter reflex arc behavior would not readily occur in a natural environment.
“During hopping an early burst can be observed in the EMG from the soleus muscle starting about 45ms after touch-down. It may be speculated that this early EMG burst is a stretch reflex response superimposed on activity from a supra-spinal origin. We hypothesized that if a stretch reflex indeed contributes to the early EMG burst, then advancing or delaying the touch-down without the subject’s knowledge should similarly advance or delay the burst. This was indeed the case when touch-down was advanced or delayed by shifting the height of a programmable platform up or down between two hops and this resulted in a correspondent shift of the early EMG burst. Our second hypothesis was that the motor cortex contributes to the first EMG burst during hopping. If so, inhibition of the motor cortex would reduce the magnitude of the burst. By applying a low-intensity magnetic stimulus it was possible to inhibit the motor cortex and this resulted in a suppression of the early EMG burst. These results suggest that sensory feedback and descending drive from the motor cortex are integrated to drive the motor neuron pool during the early EMG burst in hopping. Thus, simple reflexes work in concert with higher order structures to produce this repetitive movement.” (Received 6 October 2009; accepted after revision 6 January 2010; first published online 11 January 2010) Corresponding author J. Lundbye-Jensen: Departments of Exercise & Sport Sciences and Neuroscience & Pharmacology, University of Copenhagen, Blegdamsvej 3, 2200 Copenhagen N, Denmark.
Polysynaptic reflex arcs are distinguished from mono and oligosynaptic reflex arcs by their network connections to other neurons that are not exclusively located in the spinal cord. This description reflects the actual in vivo connectivity and functionality. In reality many – if not all – reflex arcs are polysynaptic. The intrinsic nature of monosynaptic and oligosynaptic reflex arcs are that they are embedded as specific pathways of information within a broader polysynaptic reflex arc configuration. This sets up the notions of convergence and divergence as properties of network communication.
Monosynaptic – a reflex arc with only one central processing synapse
Examples:
- Myotactic Stretch Reflex – Ia afferent to Alpha Motorneuron
- Inverse Myotactic Reflex – Ib afferent to Alpha Motoneuron
- H-Reflex – bypasses muscle spindle
- Recurrent Inhibition
Oligosynaptic – a reflex arc with two or three central processing synapses
Examples:
- Flexor Reflex
- Reciprocal Innervation
Polysynaptic – a reflex arc with four or more plus central processing synapses
Examples:
- Crossed Flexor – Extensor (withdrawal) Reflex
- Scratch Reflex
Automatic Responses
Examples:
- Postural Control
- Central Pattern Generators for Ambulation or Swimming
Control over the firing frequency of the reflex arc neurons are a basic property and is dependent on the number of central synaptic processors. Normatively, polysynaptic reflex arcs possess the property of a relatively larger number of central processors as compared to the other reflex arc configurations. This generates a greater rate of stimulation by the processors. This results in a higher degree of suppression on motor behavior.
Reflex arcs and their influence on behavior are not the only process for contributing to sensory stimulus driven motor behavior. Motor behavior can be non-reflex initiated. This motor behavior is sometimes referred to as an “automatic response”. The difference between an automatic response and a reflex arc mediated one is defined by the construction of the network connectivity within the neuronal relationships.
“Automatic movements are learned motor behaviors that are performed without conscious effort, e.g., walking an accustomed route, and tapping of the fingers while thinking about something else.” Fahn, Stanley, “Principles and Practice of Movement Disorders” 2nd Ed., 2011, Pg. 1
Observed motor behaviors that fall under the description of Automatic Responses can be caused by both sensory stimulated afferent information and/or by cerebral cortex controlled efferent information generated in anticipation of motor output. However, this type of motor behavior lacks the predictability and repeatability of motor behavior initiated and orchestrated by local spinal reflex arcs.
“The knee jerk, whether reflex or not, since it is an index of the reflex tonus of the extensor muscles, furnishes a gauge for the effect, if any, exerted by the flexion-reflex on the extensor muscles of the limb. It was said above that the extensors are not thrown into contraction by this flexion reflex. The reflex reaction may therefore either be neutral to them and leave them and their condition untouched, or it may inhibit them and depress their reflex activity, even if that activity should have at the time only the form of tonus… If the hamstring muscles (flexors of the knee) be separated from their attachments at their distal (knee) end, and then while the knee joint is passively held in approximate or full extension the flexor reflex be elicited, e.g. by electrical stimulation of the foot, the extensor muscles above the knee are easily felt by palpation to lose their tonus and relax. At the same moment the exposed and freed extensor muscles are seen to enter contraction. That is to say, the same exciting stimulus that reflexively throws the flexors into contraction interrupts reflexively the reflex tonus of the extensor muscles… This inhibition of the jerk sometimes seems to set in even before the reflex contraction of the flexors is apparent. It occurs sometimes when the stimulus is not even strong enough to evoke obvious contraction of the flexors. In the flexion-reflex therefore, the reflex excitation of the flexor muscles is accompanied by reflex inhibition of the extensor muscles both as regards their reflex tonus which is in progress when the flexor reflex is excited, and as regards their response to a stimulus (tap on tendon or muscle) that otherwise excites them” (Sherrington, 1906 & 1948).
The stimulus-response information generated and transferred to motor output control is reduced with spinal reflex arcs vs. the information generated and dispatched by higher centers. A primary system benefit of reflex arc configuration and influence on motor control is that it creates and sustains motor behaviors under conditions over time with efficiency and leads to energy conservation.
The Motor Response and Action Continuum is one where the interface of inputs that come from voluntary and non-voluntary influences overlap and support each other in an ongoing dynamic exchange. This synergistic exchange leads to motor behaviors that are intended by the individual but also able to adapt with changes in conditions and perturbations achieve a desired motor output.
“While clearly much is known about the anatomy and physiology of the motor system, there is still considerable difficulty with the concept of voluntariness. Many movements are triggered by sensory stimuli, and the physiology of this mechanism is relatively clear. However, there are certainly movements that appear to be ‘internally triggered’ and humans have the sense that they have willed the movement. The self-initiation of movement and conscious awareness of movement appear to involve mesial motor structures such as the supplementary motor area and the dorsolateral prefrontal cortex. (Dieber et al., 1999). As pointed out by Paus (2001), the mesial motor structures including the anterior cingulate cortex, in particular, is a place of convergence for motor control, homeostatic drive, emotion, and cognition. Looked at critically, the sense of voluntariness is clearly a ‘perception of consciousness’ (what can be called a quale). There is very little understanding of how this evolves.” Fahn, Stanley, “Principles and Practice of Movement Disorders” 2nd Ed., 2011, Pg. 54
Homeostatic Governance – The Autonomic Nervous System
The autonomic nervous system regulates a variety of bodily functions in order to establish and govern homeostasis and allostasis including, but not limited to; the liver, the pancreas, the kidneys, the lungs, the heart muscle, and the various glands.
“… the system of nerves that regulates the function of all innervated tissues and organs throughout the vertebrate body except striated muscle fibers; that is, the innervation of the viscera, vasculature, glands, and some other tissues.” Janig, W. “The Integrative Action of the Nervous System – Neurobiology of Homeostasis”, 2006, Pg. 13
The autonomic nervous system has three primary divisions:
(1) the sympathetic nervous system which accelerates the heart rate, constricts blood vessels, and raises blood pressure; and,
(2) the parasympathetic nervous system which slows the heart rate, increases intestinal and gland activity, and relaxes the sphincter muscles.
(3) the enteric nervous system which directly controls the gastrointestinal system.
“The essential role of the autonomic nervous system in these integrative homeostatic and allostatic programs is primarily to distribute specific signals generated in the central nervous system to the various target organs. In order to achieve overall coordination the signals need to be precisely patterned to implement appropriate reactions in each target tissue or organ.” Janig, W. “The Integrative Action of the Nervous System – Neurobiology of Homeostasis”, 2006, Pgs. 3 – 4
Allostasis (n.) – the process by which the body responds to stressors in order to regain homeostasis.
“The term “allostasis” has been coined to clarify ambiguities associated with the word ‘stress.’ Allostasis refers to the adaptive processes that maintain homeostasis through the production of mediators such as adrenalin, cortisol and other chemical messengers. These mediators of the stress response promote adaptation in the aftermath of acute stress, but they also contribute to allostatic overload, the wear and tear on the body and brain that result from being “stressed out.” This conceptual framework has created a need to know how to improve the efficiency of the adaptive response to stressors while minimizing overactivity of the same systems, since such overactivity results in many of the common diseases of modern life. This framework has also helped to demystify the biology of stress by emphasizing the protective as well as the damaging effects of the body’s attempts to cope with the challenges known as stressors.” McEwen, B., , “Stressed or stressed out: What is the difference?”, J Psychiatry Neurosci. 2005 Sep; 30(5): 315–318.
As the body is system, which by definition means interconnected, interacting, and interdependent, where many of its subsystems may be affected by a single event of another (even structural remote) system, it is understood that there is a relationship between the muscular (somatic) system and the autonomic nervous system. One area of the network can transfer information to any other area and influence that areas functionality.
“From these observations we must conclude that the visceral and the somatic afferents are connected with autonomic and somatic efferents at the segmental level of the spinal cord.” Schmidt, R.F., et al, “Fundamentals of Neurophysiology”, 2nd Edition 1978, Pg. 241.